
Endocrinal Regulation of Reproduction in Land Molluscs
Marine molluscs such as mussels, clams, winkles, squid and octopuses
provide valuable shellfish resources. On land, many slugs and snails
are agricultural pests or hosts of parasites of humans and livestock.
The future culture of shellfish and the control of pests and parasite
vectors will require a detailed knowledge of the physiology of these
species.
Most research on the endocrines of molluscs has been carried out on
mussels (Mytilus), sea hares Aplysia, garden snails (Helix) and slugs
(Limax, Arion) and the freshwater snails Helisoma and Lymnaea. Snails
and slugs are hermaphrodites, i.e. each individual produces both sperm
and eggs from a single gonad during the same breeding season, although
the animal is often a functional male before the female organs develop,
a condition known as protandrous hermaphroditism. Many of the neurons
in molluscs have very large cell bodies, and the same cells are identifiable
from their position, physiological activity or staining characteristics
in different individuals. The identity of reproductive hormones and
neuropeptide regulators of growth and reproduction is known for sure
only in freshwater snails, in which features of the anatomy (such
as the absence of tough connective tissue around the central nervous
system) make research easier than in land species. A comprehensive
scheme for freshwater species was produced by Joosse and Geraerts
in 1983. Fig. 1 shows the most widely found links in land molluscs.
Photoperiod is widely acknowledged as important in triggering sexual
maturity, but humidity, temperature and mating itself are also important.
Earlier studies relied on removal of particular organs, and manipulation
of rearing conditions, whereas more recent studies rely greatly on
immunoreactivity to possible substances.
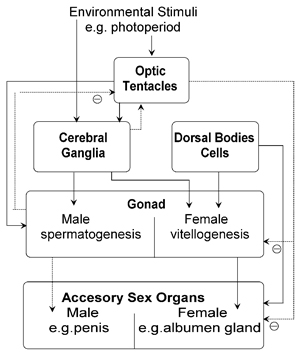
Fig. 1. Hormonal interactions in land
molluscs. Less widely established links are shown by dotted lines,
inhibitory links by a circle with a minus sign
The optic tentacles of land snails and slugs (Fig. 2) produce a hormone, the male tentacular factor (MTF) during the male phase. It stimulates sperm production, and inhibits the differentiation of the eggs and female accessory sex organs such as the albumen gland, and egg-laying. MTF is probably produced by the collar cells, which are innervated secretory cells on the side of the tentacular nerve. However, the hormonal role of the OT is controversial, not least because tentacle removal (an experimental approach that was often performed in the earlier research studies) causes severe behavioural disturbance, which may disrupt endocrine activities quite independently of any tentacular hormone.
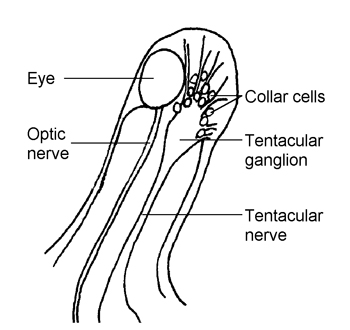
Fig. 2. Position of the collar cells
in the optic tentacle of a land snail.
Negative feedback from the gonad to the collar cells has been demonstrated
by inhibiting gonad steroid synthesis and by the direct effect of gonadial
extracts.
In the slug Limax, Sokolove et al. (1984) showed that long day photoperiods
promote the maturation of both male and female organs and the production
of a factor called Maturation Hormone (MH) from cell clusters near the
cerebral commissure (Fig. 3). Because maturation
and MH were produced in long days even if the tentacles (and eyes) were
removed, these experiments suggest (1) that the optic tentacles are
not required for maturation and (2) that daylength is perceived by an
extra-ocular pathway.
Dorsal Body Cells, DBCs
The DBCs are small groups of endocrine cells in the dorsal part of the
connective sheath surrounding the cerebral ganglia (Fig.
3). In freshwater snails they form well defined structures. They
control the development of animals from the male phase into either the
female phase or true hermaphrodites. The process involves oogenesis,
oocyte maturation, protein synthesis and growth of the female gonad
and the female accessory sex organs. The DBCs undergo seasonal changes
in size, being largest (and presumably synthesizing fastest) in the
reproductive season. They also show a 24 h cycle of packaging and secretion.
Mating triggers fine structural changes in these cells.
The chemical nature of the DBC hormone(s) is unresolved, despite much
research. Experiments suggest that DBCs either synthesize a protein
hormone and a steroid hormone, or a steroid hormone together with its
binding protein. The steroid may be ecdysone. Ecdysteroids enhance growth
and sexual maturation in the aquatic snail Biomphalaria. Ecdysone is
otherwise known only in arthropods, where it induces moulting. Insects
can only produce ecdysone from steroids in their diet, such as cholesterol,
whereas molluscs seem able to synthesize steroids.
The DBCs are non-nervous, but are innervated by neurosecretory cerebral
green cells (Ce-GCs). Transplants of part of the brain containing the
Ce-GCs into juveniles produces enlargement of the reproductive tract,
suggesting these cells hormonally stimulate the DBCs. However the DBCs
receive a complex innervation from the Ce-GCs, and the control may be
both excitatory and inhibitory. DBCs in vitro are inhibited by the peptide
FRMFamide (named from the biochemical code for the four amino-acids
it contains). FRMFamide is found in some Ce-GCs, but the stimulatory
neuroregulator is unknown.
Gonadal Steroids
The endocrine role of the gonad was established in 1954 in slugs, when
removal of the gonad was found to cause degeneration of the common reproductive
tract and the male and female accessory sex organs (the penis and the
albumen gland). Conversely, implants of gonad from mature slugs increase
cell division in male and female organs. In castrated (=gonadectomized)
slugs, exposure to long days or injection of homogenates of brain containing
MH from long day entrained slugs no longer promote development of the
reproductive tract. In freshwater snails, the development of accessory
sex organs such as the albumen gland is not regulated by the gonadal
hormones.
In vitro experiments have shown that the gonad of Helix has the enzymes
necessary to synthesize various sex steroids including testosterone
and oestradiol. In the giant African snail Achatina, testosterone synthesis
is faster during the male phase, and oestradiol synthesis is faster
during the female phase. However, no gonadal hormone has been identified
by in vivo studies. There is evidence in freshwater snails that the
Sertoli cells are the hormone producing centres in the gonad.
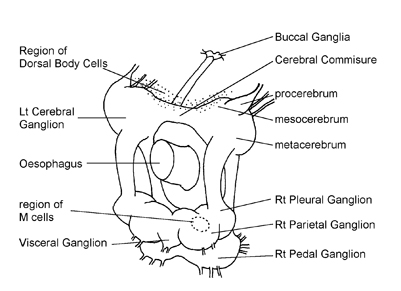
Fig. 3. The central nervous system of
a land snail, viewed from behind and from the right.


. © The Natural History Museum, London.")